CellR4 2014; 2 (1): e780
Progress Towards a Stem Cell Based Therapy for Ovarian Cancer – Proceeding of the STEMSO Conference
Topic: Uncategorized
Category: Conference Proceedings
INTRODUCTION: The Stem Cell Research Laboratory at Bradley University has an active program in research in stem cell and regenerative medicine using various adult stem cells from bone marrow, umbilical cord blood, adipose and, recently, induced pluripotent stem cells (iPS cells) and incorporating nanofiber technologies to develop potential treatments for cancer, cardiac and neurodegenerative diseases.
We are investigating the use of gene-directed cell based therapy using stem cells for treating ovarian cancer. Epithelial ovarian cancer is the most lethal of the gynecologic cancers and the fourth most common cancer in women. Although significant advances in chemotherapeutics have improved the 5-year survival rates for all stages of ovarian cancer the survival rate for metastatic ovarian cancer has not changed significantly over the last 20 to 30 years. Clearly new therapies must be developed to treat this devastating disease. Bone marrow mesenchymal stem cells (BMSC) are excellent candidates for cell-based therapies because they effectively target and persist in hard-to-reach tumor regions. BMSC are recruited to tissues following hypoxic stress or injury. BMSC may specifically target the most refractory of ovarian cancer cells, those residing within hypoxic tumor regions. Use of engineered, autologous stem cells has the potential to significantly alter the approach to treatment of ovarian cancer. This approach to stem cell gene therapy does not require bone marrow ablation, may reduce the required dose of cytotoxic therapies and may improve the sensitivity to radiotherapy for effective tumor ablation.
HYPOTHESIS: 1. BMSC preferentially migrate to ovarian cancer cells; 2. Mechanisms of migration involve secretion of specific growth factors and cytokines from ovarian cancer cells.
METHODS: Three chemotaxis models were employed to assess BMSCs migration toward ovarian cancer. 1. Tumor spheroids were generated from renilla luciferase-transduced HEY and SKOV3 ovarian cancer cells. GFP-transduced human BMSC were incubated for 24h with tumor spheroids and the spheroids examined using fluorescence microscopy for incorporation of BMSC. 2. For co-culture migration assays, BMSC (1.5 x 104 cells/ml) were labeled with 5-(6)-carboxyfluorescine diacetate, succinimidyl ester (CFDA-SE, Invitrogen) and 50 μl were added to the center of cloning rings mounted on glass coverslips. HEY, SKOV3, 2774 C10 and OCCI ovarian cancer cells, the fibroblast cell line CLL171 and the immortalized, non-tumorigenic human ovarian surface epithelium cell line, HOSE cells (0.5 ml, 1.5 x 103 cells/ml) were added to each slip, outside of the cylinder. After 3h, the cylinders were removed and the cells incubated for 48h. The cells were then labeled with 100 ng/ml 4′-6′-Diamidino-2-phenylinole dihydrochloride (DAPI, Sigma) and observed under fluorescence microscopy. Cells that migrated beyond the outer boarder of the cloning cylinder were counted. Controls included coverslips onto which BMSC were plated in cylinders without ovarian cancer cells plated outside of the cylinders. Other controls included coverslips onto which BMSC were plated in cylinders and additional, unlabeled stem cell plated outside of the cylinders. 3. For invasion assays, BMSC (1×104 cells) were plated in the upper chamber of 8μm transwell inserts, (Becton Dickenson, BD) coated with Matrigel and placed in wells containing 0, 20, 40, 80 or 100% serum-free conditioned media from either HEY or SKOV3 cells and incubated for either 24h or 48h. Positive controls included media containing 16% serum. After 24h or 48h the BMSC were removed from the upper surface of the insert membrane and the BMSC that migrated to the underside of the membrane were stained with Dif-Quik, photographed and counted.
To assess if lipid-soluble factorswere involved in BMSC chemotaxis, BMSC were plated on 8 μm transwell membranes (BD), placed in wells containing heat-inactivated (56°C for 30 min, 75°C for 15 min, or 95°C for 2 min) conditioned media from SKOV3-ip1 ovarian cancer cells and incubated for 24 h. Positive controls included media containing 16% serum. The BMSC were removed from the upper surface of the insert membrane and the BMSC that migrated to the underside of the membrane were stained, photographed and counted. For evaluation of potential cytokines that induce BMSC migration, BMSC were plated in transwell inserts with various cytokines added to serum-free media within the wells, incubated for 24h and the migrated BMSC stained and counted as above. To determine ovarian cancer cell VEGF expression, DNA was extracted from HEY, SKOV3, 2774 C10, OCCI and SKOV3-ip1 ovarian cancer cells and rtPCR performed with primers specific for VEGF-A, -B, -C, and -D. VEGF-A protein was assessed with an ELISA using conditioned media from HEY, 2774 C10, BMSC, CLL171 and HOSE cells. Migration assays as described above were used to test if increasing levels of VEGF-A (50, 100, 200 or 400 ng/mL) induced BMSC migration. BMSC co-culture migration assays were performed as above using CLL171 fibroblast cells transduced to overexpress VEGF and non-transduced CLL171 cells served as a negative control. To determine if pre-treating BMSCs with VEGF and ovarian cancer conditioned media would enhance BMSC migration toward ovarian cancer BMSCs were treated for 24 hours with HEY cell conditioned medium with or without VEGF-165 (150 ng/ml). Following treatment BMSCs were plated in 8 µm transwell inserts, placed in wells containing HEY cell conditioned media and assayed using the migration assay detailed above. Data were analyzed using Prism for Macintosh (GraphPad Software, San Diego, CA). Statistical differences among treatment groups for invasion assays were assessed using a one-way analysis of variance (ANOVA) followed by Dunnett’s post-hoc testing when indicated. Unpaired t-tests were performed to assess differences in stem cell migration between groups with and without ovarian cancer cells at 24 and 48 hours. Probability values <0.05 were considered statistically significant.
RESULTS: For tumor spheroid migration assays, BMSC were observed surrounding and within tumor spheroids following 24h (see Figure 1).
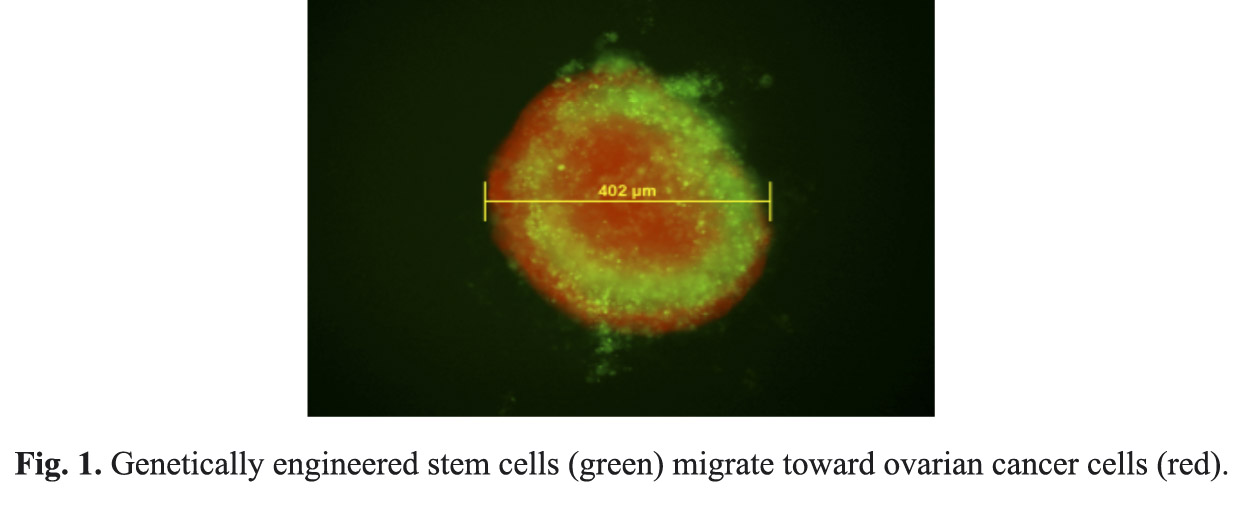
In co-culture assays, there was significant migration of BMSC toward all ovarian cancer cell lines at 24 and 48 h (p < 0.0001). Co-culture with CLL171 or HOSE cells did not induce BMSC migration. For the invasion assays, 40, 80 and 100% conditioned media from the ovarian cancer cell lines HEY and SKOV3 significantly induced BMSC invasion compared to serum-free media (p < 0.001). For heat-inactivation experiments to test for lipid-soluble factors that may stimulate BMSC migration, there was a significant reduction in migration for heat-inactivated conditioned media relative to non-heat-inactivated conditioned media (56°C, p < 0.05, 75°C and 95°C, p < 0.001). Multiple cytokines stimulated BMSC migration. Relative to serum-free control, VEGF-165 exhibited a 29% increase, VEGF-C, 22%, VEGF-121, 26%, mVEGF 15%, FGF, 21%, Wnt-1, 32%, SDF-1A 27%, HGF-1 16% and TGF-beta1, 19%. Whereas IGF-1, ANG-1, ANG-1, GM-CFF, IL-3 and ET-1 all exhibited less than 1% increase in migration relative to the serum-free control. In rtPCR experiments, VEGF gene expression was demonstrated in all ovarian cancer cell lines. VEGF was detected with ELISA in conditioned media from 2774 C10 (3249 ± 242 pg/mL), SKOV3-iP1 (612.8 ± 102.3), HEY (1171.7 ± 2.5). The control cell line CLL1781 exhibited a much lower concentration of VEGF, (36.7 + 1.4), whereas HOSE cells had non-detectable levels of VEGF. For dose-response migration assays, all concentrations of VEGF tested resulted in significant migration of BMSC compared to migration observed with serum-free media (p < 0.001). In co-culture migration assays with CLL171 cells overexpressing VEGF, BMSC migration was significantly greater at both 24 and 48h compared to migration of BMSC with non-transduced CLL171 cells (for 24h, p < 0.01, for 48 h, p < 0.05). In migration assays we found that pre-treatment of BMSCs with cancer cell conditioned media and VEGF-165 significantly enhanced BMSC migration toward ovarian cancer cell conditioned media (conditioned media, p < 0.01, VEGF-165, p < 0.05) relative to untreated BMSCs.
CONCLUSIONS: Our results suggest that BMSCs migrate toward ovarian cancer cells and that soluble factors, including VEGF mediate this migration. We have also confirmed that BMSC migration toward ovarian cancer cells can be enhanced by pretreating cells with either VEGF or ovarian cancer cell conditioned media. Autologous, engineered BMSCs may specifically target the most refractory of ovarian cancer cells, those residing within hypoxic tumor regions and significantly improve the approach to treating ovarian cancer. A stem cell based therapy does not require bone marrow ablation, may reduce the required dose of cytotoxic therapies and may improve the sensitivity to radiotherapy for effective tumor ablation. At the very least a stem cell based therapy may become another addition to the growing armamentarium for the treatment of more refractory or more advanced cases of ovarian cancer.
To cite this article
Progress Towards a Stem Cell Based Therapy for Ovarian Cancer – Proceeding of the STEMSO Conference
CellR4 2014; 2 (1): e780
Publication History
Published online: 05 Feb 2014

This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.