CellR4 2014; 2 (3): e963
Magnetic Resonance Imaging of Stem Cell Transplantation in Injured Mouse Spinal Cord
Category: Original Articles
Abstract
BACKGROUND: Several types of cell-mediated treatments have been studied in order to establish their potential usefulness in promoting neural tissue regeneration and repair after spinal cord injury (SCI). We recently isolated a subclass of mouse neural progenitors, capable of surviving a powerful ischemia insult, and their differentiation yield mostly neurons compared to regular neural stem cells (NSCs). One of the main difficulties in analyzing the effects of transplanted cells during the time is the distribution evaluation of injected cells and their target organ localization.
MATERIALS AND METHODS: We labeled NSCs with Iron Oxide paramagnetic nanoparticles (Endorem®), stem cell features and differentiation capabilities of iron-labeled cells were investigated by immunofluorescence. Iron-labeled cells were injected in tail vein and their distribution was tracked by Magnetic Resonance Imaging (MRI) and demonstrated histochemically.
RESULTS AND DISCUSSION: We observed that the increase in number of iron-positive cells during the labeling procedure was proportional to the iron concentration in the medium. A higher labeling was observed when Protamine sulfate (PrS) was used as carries. Stem cell features and differentiation capabilities were unaffected by iron-labelling. MRI visualization showed that labeled NSCs were accumulated at the lesion site, differently no labeling was detected in the injured cord when free iron nanoparticles or dead labeled cells were administered. Moreover, labeled NSCs promoted hind limb recovery of function through the experimental period.
CONCLUSIONS: Our study shows that NSCs can be efficiently labeled with dextran-coated paramagnetic nanoparticleswithout significant alteration of their physiological and functional features. MRI may result useful in the study of stem cell localization at the lesion site in murine models of SCI
L. Ottobrini and S. Carelli contributed equally to this work.
INTRODUCTION
In traumatic spinal cord injury (SCI), destruction of nerve fibers that carry motor signals from the brain to the torso and limbs leads to muscle paralysis and often also to loss of sensation. The tissue damage caused by SCI derives from two distinct phases. The primary lesion is caused by the mechanical impact, and it is followed by the secondary lesion, that is the consequence of multiple cascades of events triggered by the ischemic insult, and the vigorous inflammatory response. These latter events eventual lead to further tissue destruction with extensive neuronal and glial cell loss 1 , 2 , 3 , 4 , 5. Several types of cell-mediated treatments have been studied in order to establish their potential usefulness in promoting neural tissue regeneration and repair after SCI. We recently reported that adult mice neural stem cells (NSCs), applied i.v. following experimental spinal cord injury, promoted hind limb recovery of function but with a reduced survival 6. Differently, the recovery of function was smaller and with no survival of transplanted cells at lesion site, when we transplanted embryonic (ESCs). In addition we observed the colonization of the intact lumbosacral region of the cord. The recovery of function was associated with reduced invasion of the injured cord by inflammatory cells 7 , 8.
A similar effect was observed when term amniotic fluid cells were used 9. Thus it is apparent from our experience that adult NSCs are capable of reaching the lesion site where they promote series of positive events but their fate is unclear. Thus the development of adequate monitoring techniques for assessing the fate of transplanted cells is desirable.
One of the main difficulties in analyzing the effects of cell treatments is that the duration of evaluation of their efficacy on functional recovery can vary, ranging from a few weeks to several months after injury and transplantation. Important variables such as the distribution of injected cells, their target organ localization, and their survival, proliferation and differentiation are normally assessed at sacrifice, with no data being collected during the observation period. The development of molecular imaging strategies could help to overcome these limitations, by providing refined methods for the in vivo visualization and study, in real time, of biological events associated with the use of cell-mediated therapies.
Indeed, molecular and cell imaging techniques could provide information about the homing and fate of injected stem cells, and they could also provide information regarding the effect of pharmacological treatments on the fate and function of applied cells. Specific cell populations can be labeled and visualized in vivo through widely used methods such as magnetic resonance imaging (MRI), emission tomography (PET/SPECT) and, only for animal models, bioluminescence (BLI) and fluorescence imaging (FLI). Cell imaging gives insight into some of the fundamental aspects of stem cell therapy, allowing the in vivo assessment of distribution and fate of appropriately labeled cell populations both in the preclinical and in the clinical settings 10 , 11. In protocols based on the use of stem cells, indeed, in vivo imaging can provide a novel approach for studying stem cell homing and differentiation into the tissue of interest.
Two main strategies have been proposed for cell labeling: direct and indirect. Direct labeling involves the internalization of an agent that can be visualized in the specific cell population under examination. In this case, however, proliferation leads to marker decay because of the symmetric segregation with mitotic divisions. Various reports on stem cell labeling with different types of paramagnetic nanoparticles 12 , 13, 14 , 15 , 16 , 17 , 18 , 19 , 20 , 21 and radiotracers 22 , 23 suggest that it is possible, also, using clinically approved materials, to track these cells without altering their physiological features. Indirect labeling, on the other hand, involves the stable introduction of a reporter gene, whose persistence and expression are not influenced by cell proliferation. For the moment, however, these strategies look destined to remain limited to pre-clinical settings.
MRI has been shown to be the most attractive technique for in vivo imaging-based evaluation of cell-mediated treatments. Indeed, it can produce images with high resolution and with high contrast for soft tissues; it is also safe, since it does not use ionizing radiation. Furthermore, the existence of similar instruments both for small-animal and for clinical use, gives it great translational potential. Finally, several contrast agents for cell labeling and visualization by MRI have been produced and approved for clinical use (i.e. Resovist®, Endorem®). Some studies of MRI evaluation of stem cell-mediated treatments in both rodent 24 , 25 , 26 and non-human primate 27 SCI models have been reported.
We herein describe the use of a direct labeling strategy (involving paramagnetic iron oxide nanoparticles) that allows visualization by MRI of murine adult SC localization and persistence at the target organ in a murine model of SCI.
METHODS
Murine NSC culture
Murine SCs isolated from the subventricular zone (SVZ) of CD1 mice 28 6h after sacrifice were cultured in DMEM/F12 medium (supplemented with 2mM L-glutamine, 0.6% glucose, 9.6 g/mL putrescine, 6.3 ng/ml progesterone, 5.3 ng/ml sodium selenite, 0.025 mg/mL insulin, 0.1 mg/mL transferrin and 2 μg/mL heparin, control medium) and with adding of FGF2 (20 ng/mL) and EGF (20 ng/mL) 29. The medium was changed every 3-4 days, until the neurospheres appears. At each passage, spheres were centrifuged and dissociated in the culture medium and then replated at the concentration of 5×105 cells/mL 30.
Labeling of NSCs with superparamagnetic iron oxide nanoparticles (SPIOs)
Murine SCs were freshly plated early in the morning. Six hours later, cells were labeled by adding different amounts of SPIOs (0 – 100 – 200 – 400 μg Fe/ml final concentration Endorem®, Guerbet, Roissy CdG Cedex, France; stock solution 11.2 mg Fe/ml) in the culture medium and incubated for 24, 48 or 72 hours in the presence of different transfection agents, namely poly-L-lysine hydrobromide (PLL, Sigma-Aldrich, St. Louis, MO, USA; stock solution 2.4 mg/ml), polybrene (PB, Sigma-Aldrich; stock solution 10 mg/ml, used at 10 μg/ml) and protamine sulfate (PrS, Sigma-Aldrich; stock solution 10 mg/ml). PLL and PrS were tested at different ratios (Fe/PLL 1:0.03, 1:0.06 and 1:0.09 and Fe/PrS 1:0.025 and 1:0.05). Cells were washed with phosphate-buffered saline (PBS) supplemented with heparin (1 U/ml). After 24, 48 and 72hours ofincubation, cells were counted to analyze viability bythestandard trypan blue exclusion testand then washed. Morphology (Perl’s staining), iron content, and relaxation time reduction were analyzed to evaluate labeling efficiency and the in vivo detectability by MRI of the cells. Part of the labeled cells were also plated and cultured for a further 5 or 7 days to allow analysis of their features after labeling.
Labelling evaluation
Iron content evaluation
After incubation with paramagnetic nanoparticles, Prussian blue staining (Perl’s acid ferrocyanide) was used to detect iron in cell cultures. 150,000 labeled NSCs were dissociated to a single-cell suspension, harvested on microscopy glasses using cytospin, and fixed with Cervix fixative (Avantor Perfomance materials, Phillipsburg, NJ, USA). Prussian blue staining was performed by incubation in a 1:1 mixture of 2% HCl (BDH Laboratory Supplies, Dubai, UAE) and 2% potassium ferrocyanide (Merck, Whitehouse Station, NJ, USA) for 45 minutes at room temperature (24°C). Cell membranes were counterstained with Gram’s safranine solution (Merck, Whitehouse Station, NJ, USA). Labeling efficiency for analyzed by counting a minimum of 7 non-overlapping fields for each sample. Data were expressed as mean percentage of Prussian blue-positive cells (over total counted cells). Iron content per cell was moreover evaluated by spectrophotometric analysis 31.
Permeabilization Shift Assay
After 24 h labeling with or without 200 g of SPIOs and PrS, permeabilization shift assay was performed as previously described 32.
Phenotypic differentiation of iron NSCs
NSCs ability to differentiate into the 3 typical neural cell lineages were retained after SPIOs labeling through immunofluorescence analysis for GFAP (marker of astrocytes), Gal-C (marker of oligodendrocytes) and beta-tubulin III (neuronal marker) expression, after 7 days of in vitro differentiation.
Differentiation of NSCs was achieved after plating the dissociated stem cells at the density of 6×104 cells/well in presence of adhesion molecules (MatrigelTM) and bFGF (10 ng/ml) for 48 h. The growth factor was removed during the following 5 days and fetal bovine serum (1% vol/vol) was added to the medium 33. Differentiated cells were fixed in paraformaldehyde (PFA) 4% for 10 minutes. Cells were then washed in PBS before and after the treatment with PFA and stored in PBS at 4°C.
Spinal cord injury
Animals
Adult CD1 mice (28-30 g; Charles River) were used. A standard dry diet and water were available ad libitum. All experimental protocols were approved by the Review Committee of the University of Milan and met the Italian guidelines for the treatment of laboratory animals, which comply with the European Communities Directive of November 1986 (86/609/EEC). Animals were kept in standard conditions (22 ± 2° C, 65% humidity, and artificial light between 08:00 a.m. to 08:00 p.m.) for at least 3 days prior to the experiments.
Spinal cord injury
Moderate spinal cord injury was induced using an Infinite Horizon (IH) Impactor (Precision Systems and Instrumentation, Kentucky, USA). The surgical procedures were performed as described previously 34 , 35. Animals were anesthetized by intraperitoneal injection (10 ml/kg) of 4% chloral hydrate (Sigma-Aldrich). The anesthetized mice were submitted to laminectomy at vertebral level T9. The lateral processes of T8 and T10 were cleared of muscle to allow stabilization of the vertebral column using forceps attached to the impactor-clamping platform, and a contusion injury was induced on the exposed cord using the impactor device as described 36 , 37. Muscle and skin were sutured after injury by using absorbable suture threads (Etichon, Cincinnati, Ohio, USA). To prevent post-operative sepsis, all mice were treated with 100 ml of ampicillin (100 mg/kg, Farmitalia, Milan, Italy), injected subcutaneously, and 1 ml of physiological saline solution. Before awakening, they were treated with buprenorphine (a single injection of buprenorphine at dosage of 0.03 mg/kg) (Sigma-Aldrich) for pain relief, and penicillin G (10,000 U/kg; Sigma-Aldrich) as an antimicrobial agent.
Experimental groups and transplantation
After incubation in SPIO-containing medium, cells were washed in PBS supplemented with heparin (1 U/ml) and viable cells were counted by trypan blue exclusion.
The animals were divided into four experimental groups:
1) spinal cord injured CD1 mice in which SPIO-labeled NSCs were injected directly into the spinal cord (positive controls) (n=3);
2) spinal cord injured CD1 mice injected with SPIO-labeled NSCs into the tail vein (n=8 );
3) spinal cord injured CD1 mice injected with PBS or non-labeled cells (negative controls) (n=3);
4) spinal cord injured CD1 mice injected with dead SPIO-labeled NSCs into the tail vein (n=3).
In the intraspinal injections, immediately after the lesion we performed two stereotaxic injections of 75 x 103 cells (or the equivalent volume of PBS) 2 mm rostrally and caudally at the site of the injury. In the intravenous injections, 1 x 106 cells were injected into the tail vein in 2 injections immediately after the lesion and 30 minutes later (5 x 105 cells/injection).
Locomotor recovery assessment
Locomotor function and hind limb recovery after contusion were evaluated in blind with the open field test according to the Basso mouse rating scale as previously described ([ii]). Evaluations were performed 24h after injury and once a week for the next 4 weeks.
In vivo MRI
Magnetic resonance imaging was performed using a 7T MR scanner (Bruker). MRI analysis was longitudinally performed from 1 week up to 5 weeks after transplantation in both intraspinally and intravenously injected animals.
Anesthetized animals (isofluorane gas anesthesia) were positioned supine on the animal bed and inserted in the radiofrequency coil (38 mm i.d.) inside the magnet. Scout images were acquired for correct positioning of the region of interest. Different MR sequences were tried in order to optimize spinal cord visualization and signal contrast:
- Spin echo (MSME) sequence (TR/TE: 1200/12 ms; 2 averages; acquisition time: 5′ 7”),
- Gradient echo (FLASH) sequence (TR/TE: 1200/10 ms; 2 averages; acquisition time: 5′ 7” + resp. triggering).
- Fast spin-echo (RARE) sequence (TR/TEeff: 2000/56 ms; 8 averages; rare factor=8; acquisition time: 4’16’’).
All animals were acquired in coronal (5 slices of 0.7 mm thickness; FOV 4×4 and spatial resolution of 156 µm) and axial orientations (12 slices of 0.7 mm thickness; FOV 3×3 and spatial resolution of 117 µm).
Ex vivo MRI
Collected spinal cords were positioned inside needle caps in PBS, placed on the animal bed and inserted in the radiofrequency coil inside the magnet. To visualize them, FLASH 3D sequences were used with the following parameters: FOV=2*2*1.6 cm; matrix 256*128*64; spatial resolution of 78 µm; TR=30 ms; TE=6.2 ms; flip angle=15; 30 averages; scan time=2h2′.
Histology
Spinal cord sections were analyzed by optical microscopy at 5X, 20X and 40X original magnification after Prussian blue staining and counterstained with Gram’s safranine solution (Merck) to confirm the homing of labeled NSCs.
Longitudinal sections (3-4 μm) were cut and stained with hematoxylin and eosin (H&E) for the morphological evaluation. The deparaffinized sections were immunohistochemically assessed using the Genomix i-6000 automated staining system (BioGenex, San Ramon, CA, USA). Heat-induced antigen unmasking was performed using either EDTA (0.05 M, pH 8) or citrate (0.01 M pH 6) solutions in a thermostatic bath for 35 min. Then slides were incubated with monoclonal anti-Macrophages/Monocytes antibody (clone MOMA-2, MAB1852 Millipore KGaA, Darmstadt, Germany). Reactions were revealed using the Novolink Max polymer detection system (Leica Biosystems), in accordance with the kit manufacturer’s instructions, with the polymer for 30 min and with DAB for 8 min. Positive cells were counted in five fields of high magnificence (40X). The values obtained in the five fields were averaged for each section.
Statistical analysis
Statistical analysis was performed using a Student’s t test and ANOVA. Significance was accepted at p < 0.05.
RESULTS
NSCs labeling with SPIOs
The extent of cell labeling was assessed after SPIOs exposure to the Endorem® (Guerbet) with or without co-incubation with different carriers, namely PLL, PB and PrS, that form complexes with the SPIOs. Labeling efficiency was calculated by assessing the percentage of cells positive to specific Perl’s iron staining, while the viability of labelled cells was evaluated by standard trypan blue exclusion test.
Murine adult NSCs were incubated for 24h with different amounts of SPIOs (25-50-100-200-400 μg Fe/ml, Endorem®) in the presence or absence of different carriers. After Perl’s staining, it was observed that the extent of cell labeling with SPIOs was found to be dependent on the concentration of iron in the culture medium, and on the presence or absence of carriers (Figure 1 A,B). In particular, different carriers had different effects on cell labeling efficiency and cell viability and incubation of NSCs with SPIOs alone or in the presence of PLL or PB as carriers produced inefficient cell labeling at all iron concentrations. Furthermore, increasing carrier concentration failed to improve labeling efficiency while cell viability was reduced (data not shown). Conversely, the use of PrS (ratio Fe/PrS 1:0.025) as carrier greatly improved cell labeling and 96.01±0.3% of positive cells were obtained with 200 μg Fe/ml (Figure 1 A, B). Lower iron concentrations (25-50-100 μg Fe/ml) were soon discarded due to their insufficient labeling efficiency. On the contrary, higher iron concentrations (400 μg Fe/ml) resulted in evident toxicity and lower cell viability (Figure 1C). Different PrS/iron ratios and different incubation time were tested, but no improvements in cell labeling was observed and this was accompanied by significant reduction of cell viability (data not shown).
The assessment of intracellular iron content assessment 38 confirmed the higher labeling efficiency when PrS was used. NSCs cultured in the presence of 200 μg Fe/mL for 24 hours incorporated 130±28.28 pg Fe/cell in the presence of PrS as opposed to 11.26±3.1 pg Fe/cell when in presence of PLL. Iron content was not evaluated in presence of PB due to the excessive cell toxicity of this carrier. Non-labeled control cells presented a very low intracellular iron level (3.56±2.6 pg Fe/cell).
Compared to control cells, the use of PrS as carrier did not significantly affect cell viability or proliferation capacity when NSCs were incubated for 24h with 200 μg Fe/ml (Figure 1C).
These data strongly suggest that 200 μg Fe/ml for 24 hours in presence of PrS is the optimal cell labeling condition for NSCs.
A permeabilization shift assay was used for the evaluation of the ratio between the iron internalized and that surface-absorbed in unlabeled controls and labeled cells with 200 μg Fe/mL in presence of PrS (Figure 1D). A clear permeabilization-dependent shift can be observed exclusively in the FACS profile of SPIO labelled cells (Figure 1D). This shows that iron internalization occurs in at least 99.3% of cells, and there is no surface absorption of the nanoparticles in the cell membrane.
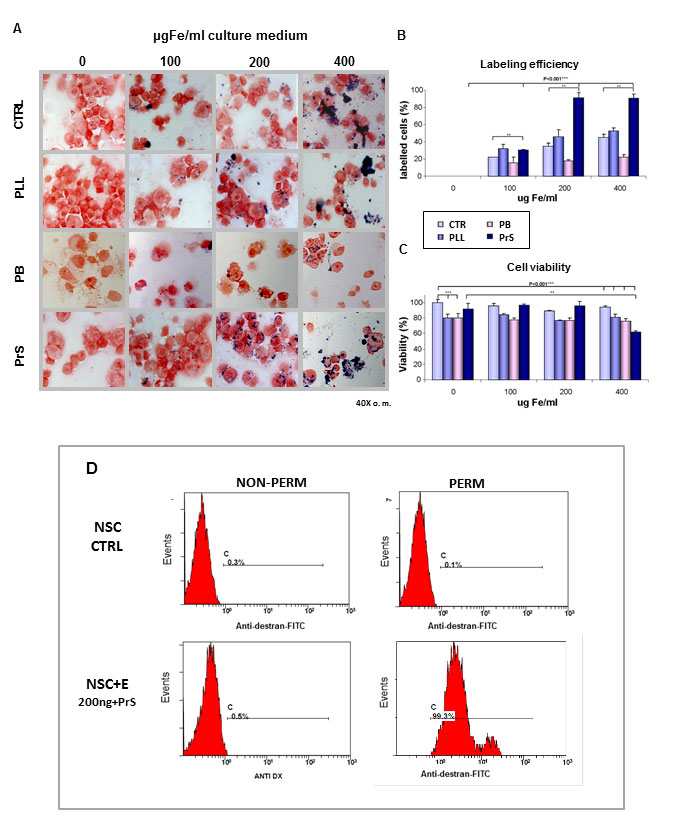
Figure 1. Efficiency of NSCs labeling. Adult NSCs isolated from the subventricular zone of CD1 mice were labeled with different amounts of SPIOs (0, 100, 200, 400 μg/mL Fe) in the presence or absence of different transfection agents, namely poly-L-Lysine (PLL), polybrene (PB), protamine sulfate (PrS) for 24h (A). Perl’s staining of labeled cells showed that cell iron content increased in a dose dependent manner of SPIOs added to the culture medium. The effect was also dependent upon the addiction of transfectants. In presence of the carrier PrS, the percentage of iron-positive cells increased proportionally to the iron concentration. In particular, we obtained 96.01± 0.3% iron-positive cells in samples incubated with 200 μg Fe/ml for 24h in presence of PrS (p < 0.001, n=7 compared to controls (B). A higher concentration of iron (400 μg Fe/ml) promoted a larger increase in iron content per cell, but cell viability was strongly affected (p < 0.001, n=20). The percentage of viable cells also decreased when carriers PLL and PB were used (p < 0.001, n=9 for PLL and n=2 for PB). Differently, PrS did not influence cell viability labelling when used for labelling cells with 100 and 200 μg/mL SPIOs. Cell toxicity was observed whit all the carriers when higher SPIO concentrations were used (C). Iron labelled cells reached 99.3% as show by the dextrane-positive FACS profile when Prs was used (D).
Unmodified NSCs phenotype after iron labeling
The maintenance of typical NSC marker expression before and after differentiation and the ability to generate neurospheres was tested. NSCs were washed and replated the neurosphere assay medium after labeling with SPIOs (200 μg Fe/ml with PrS ratio 1:0.025 for 24h of incubation). Within 5 days these cells formed neurospheres with the same efficiency observed in control cultures [172± 9.07 spheres/well (control NSCs) vs 168± 11.21 spheres/well (labelled NSCs)].
Following differentiation, iron-labeled NSCs showed the classical GFAP, GAL-c and typical and beta-tubulin III positive staining suggesting that iron labeling do not modify specific cell features (Figure 2). Also the extent of such antigen specificity was comparable to controls [beta-TUB III 32.75±2.71% (control NSCs) vs 31.86±3.21% (labelled NSCs); 65.24±3.45% (control NSCs) vs 66.45± 2.89% (labelled NSCs); Gal-C 6.82±3.12% (control NSCs) vs 6.97±2.34% (labelled NSCs)].

Figure 2. Stability of NSCs phenotype. NSC labeling with SPIOs did not affect the typical phenotypic differentiation. NSCs labeled with Endorem® maintained the ability to form new neurospheres within 5 days in culture when replated soon after procedure labeling (A). Differentiated NSCs, after being labeled with SPIOs (200 μg Fe/ml and PrS ratio Fe/PrS 1:0.025), were positive to anti β-tubulin III antibody (B, green), to anti-GFAP antibody (C green), and to anti-Gal-C antibody (D, green). Nuclei were stained with DAPI (blue).
In vivo MRI after intra-spinal administration of labeled SCs
In vivo tracking, by MRI, oflabeled NSCs directly implanted into the spinal cord of injured mice was performed. 1.5 x 105 labeled NSCs were injected intra-spinally within 60 minutes of injury. MRI, performed one week after cell transplantation, showed, on FLASH images, a strong and well defined region of signal void in the spinal cord at the lesion site in all intra-spinally injected mice (n=3) (Figure 3A). The area of signal loss was persistent throughout the experimental period (data not shown) appeared to diminish slightly over time, but a strong hypointensity persisted for at least 5 weeks (data not shown) suggesting the persistence of transplanted cells.
The presence of labeled NSCs at the site of injection in the lesion cord was also shown by histology of the injured spinal cord. Several cells were positive to Perl’s staining, thus confirming the presence of iron-labeled cells at the site of injury (Figure 3B).

Figure 3. MRI-mediated in vivo tracking of intra-spinally injected labeled NSCs. In vivo MRI (gradient-echo, FLASH) allowed iron signal detection in the injured spinal cord of CD1 mice after the intraspinal injection of SPIO-labeled NSCs (200 μg Fe/mL+PrS for 24h). MRI, using a Bruker 7T Pharmascan, was performed once a week for 5 weeks, beginning 7 days after the transplant. SPIO-labeled cells were detectable soon after injection. The white dotted circle indicates the lesion site while the yellow arrow points to the SPIO-labeled NSCs (A). The images refer to one of three animals. The histological analysis, after Perl’s staining of spinal cord lesion site shows clearly the presence of SPIO-labeled cells at the margin of the lesion (B).
In vivo MRI NSCs localization after intra-venous administration
Localization of NSCs at the lesion site after the two intravenous injections of 106 SPIO-labeled NSCs (5 x 105 cells/injection) through the tail vein of injured animals was visualized by performing serial MRI scans once a week, starting 1 week after spinal cord injury and cell injection, up to 5 weeks. A hypointense signal was detectable since 1 week after cell injection (n=8). The signal was characterized by a clear region of hypointensity over the entire spinal cord section at the lesion epicenter, that progressively was restricted to the dorsal portion of the spine when looking further away from the lesion (Figure 4B). Figure 4A compares the MRI signals obtained with different acquisition sequences in injured control animals or after i.v. administrations of SPIO-labelled cells. MSME images showed some hypointensity, that was better-defined as a dark region by the FLASH and RARE images. RARE acquisitions, like MSME, are spin-echo sequences able preserve anatomical detail, but, being strongly T2-weighted (in comparison with MSME), they better highlighted the hypointensity caused by the accumulated iron-labeled cells at the site of injury. Another feature observed was the shrinkage of the spinal cord matter at the lesion site; this appeared on FLASH and RARE images as a hyperintense signal, probably due to spinal liquor filling empty spaces in the spinal canal. The presence of liquor at the lesion site is in line with the very high signal found on strongly T2-weighted RARE images. Spinal cord shrinkage at the lesion site and liquor accumulation was a constant finding in all animals with some differences among the two animal groups probably secondary to the effect of the cellular treatment. Spinal cord shrinkage at injury site was also a clear finding by the ex vivo MRI analysis, performed on isolated spinal cords (Figure 4B). Axial slices were obtained from the 3D data set and confirmed the hypointense signal located along the dorsal portion of the spinal cord. Central slices showed hypointensity covering almost the whole cord diameter, and the higher resolution of ex vivo MRI made possible to distinguish a granular pattern of hypointensity.
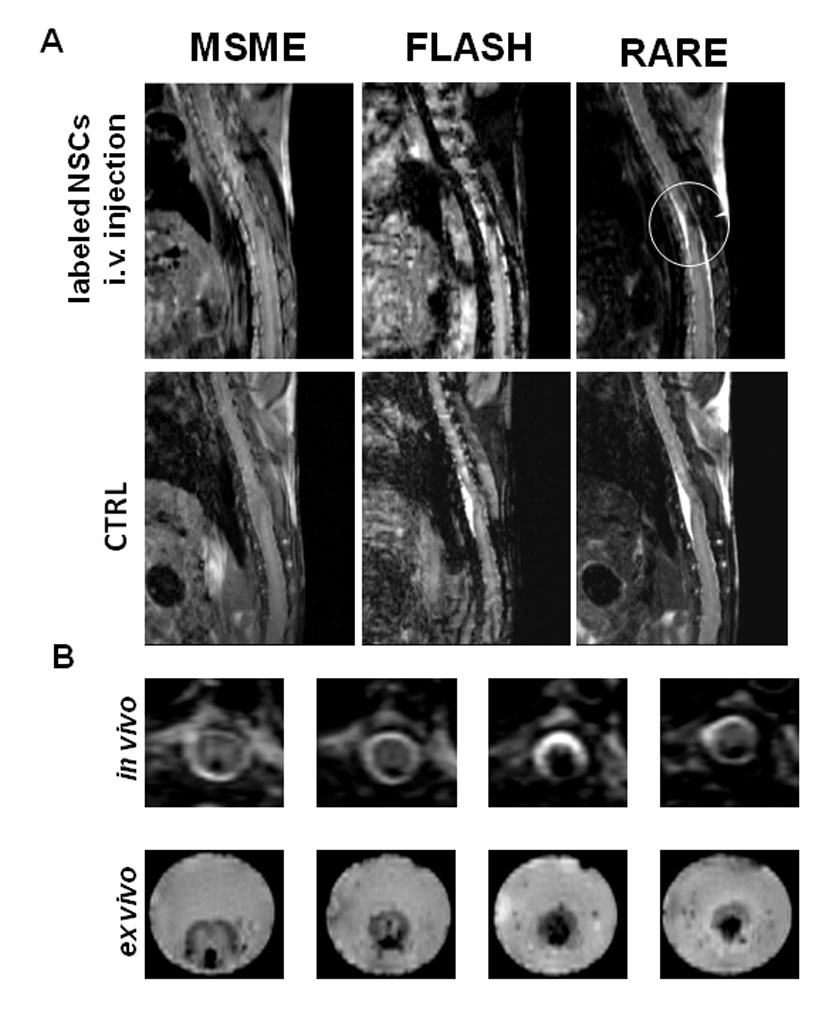
Figure 4. MRI In vivo and ex vivo tracking of intravenously injected labeled NSCs. Different MRI sequences (MSME, FLASH and RARE) were tested the detection of intravenously injected SPIO-labelled NSCs. MR imaging, using a Bruker 7T Pharmascan, was at first performed 7 days after transplant and was repeated once a week for the following 5 weeks. Mice were positioned supine and MSME, FLASH and RARE sequences were acquired. All the acquisition sequences highlighted the hypointense signal due to the presence of iron-labelled NSCs at the lesion site (yellow dotted circle). RARE sequences showed both a hypointense signal due to labelled cells and a hyperintense signal produced by the change in the spinal cord/liquor ratio at the lesion site. The traumatic lesion has caused a reduction in size of the intact cord (A). Ex vivo MRI (lower line) of the injured spinal cords collected 6 weeks after transplantation revealed the same hypointense signal observed in the in vivo MR images (upper line) at the lesion site (B). The images all refer to one of eight animals. The data are highly consistent and repeatable form animal to animal.
Mice were acquired by MRI once a week from the 1st to the 5th week after intravenous cell administrations. Figure 5A shows the kinetic of the hypointense signal at the lesion site during this time. The signal persists in injured animals injected with SPIO-labeled cells. Differently, only a weak nonspecific signal void was observed with dead-labelled cells in the first two weeks and no further signal void was detectable later. The histological analysis carried out five weeks after lesion on sagittal sections confirmed the presence of Perl’s staining positive SPIO-labeled cells at the lesion site and in the dorsal portion of the spinal cord, in proximity to the lesion (Figure 5B). Conversely, no iron labeled cells were found at the lesion site of animals intravenously injected with both SPIO-labelled dead cells (Figure 5B) and control cells (data not shown).
Hind limb locomotor recovery was assessed in injured mice treated with saline, or NSCs, or SPIO-labeled NSCs, or dead SPIO-labeled NSCs. The time course is reported in Figure 5C, where it is evident that both unmodified NSCs and SPIO-labeled NSCs promote hind limb recovery of function in a comparable manner through the experimental period, and that dead NSCs were not better then saline as expected. This is a qualitative and quantitative confirmation of previously reported effects of adult NScs on acute spinal cord injury 39.

Figure 5. MRI dynamic imaging of NSCs persistence at the lesion site. Labeled NSCs, injected intravenously (immediately after the injury and 30 minutes later), were detected at the lesion site a week after injection and persisted for the following weeks. The hypointense signal decreased only slightly with time, demonstrating that SPIO-labeled SCs localized at the lesion site remained in situ throughout the experimental period (A). Histological Prussian blue analysis of the same cord described in panel A shows the presence of iron-labeled cells in the lesion spinal cord, and this is a confirmation of the hypointense pattern revealed by in vivo MRI (B). The hind limb locomotor recovery in all animal groups was tested according to the Basso mouse rating scale once a week for the entire experimental period and it is shown that SPIO labeling does not negatively modify NSC-promoted enhancement of hind limb recovery of function. Dead SPIO-labelled cells on the other hand failed to modify the extent of recovery and the outcome was comparable to the control injected with PBS (C). The groups were randomized, the analysis was performed in double blind fashion. Values represent average ± SEM. We determined the statistical differences by means of ANOVA test followed by Tukey’s post-test. ***p < 0.001; **p < 0.01 vs Saline; °°°p < 0.001; °°p < 0.01 vs dead NSCs.
We had previously reported that NSCs reparatory action was accompanied by antinflammatory effects 40. Here we show that also, after iron labeling, NSCs counteract the invasion of the injured spinal cord by macrophages. Figure 6 shows that the number of ED-1-positive cells (macrophages) is significantly reduced in the site of lesion in animals treated with SPIO-labeled NSCs at one week after injury.

Figure 6. Counteraction of MOMA-2 positive cell invasion of spinal cord. We had previously shown that NSCs reduce the extent of macrophages infiltration into the injured cord. Here we show that such ability is maintained also when NSCs were labelled with iron. The images are representative for two mice, but similar images were obtained for at least three animals per group. For the quantification we considered sections from at least 3 animals per group. Values represent average ± SEM. We determined the statistical differences by means of t-test.
DISCUSSION
Cell-based therapy is seen as a promising strategy for the treatment of CNS neurodegenerative diseases. We have conducted a preclinical study on the in vivo fate monitoring of adult NSCs transplanted into the traumatically injured spinal cord 41.
The preclinical assessment of the efficacy and safety of these novel therapeutic approaches could move them closer to the clinical testing phase.
Important questions about the molecular mechanisms underlying recovery after cell-mediated treatments, the optimal treatment protocol (particularly with regard to the type and amount of cells to use, the therapeutic window, and their fate) must first be answered. Therefore, molecular and cell imaging can clearly provide a fundamental contribution in the preclinical and possibly, in the future, also in the clinical testing of these procedures. These techniques could allow early and accurate evaluation of important variables related to the final efficacy of the treatment. For example, the study of cell homing to target tissues could open the way for dynamic evaluation of the relationship between therapeutic cell localization at the lesion site and motor recovery, and for the development of personalized treatment schedules. Indeed, non-invasive, imaging-based methods have recently been developed to track, in vivo, transplanted cells that had first been appropriately labeled 42. These imaging techniques make it possible to monitor the distribution, migration and localization of cells introduced into the living model. The ability to in vivo monitor the location of cells would allow to non-invasively asses if therapeutic cells once injected intravenously are able to migrate to the target tissue and how long can persist at the lesion site.
Cell labeling can be performed using two different strategies: direct and indirect. Given the difficulties involved in the genetic manipulation of cells for clinical use, this study focused on the use of a direct labeling procedure based on clinically approved reagents. There already exist several clinical diagnostic procedures (for occult infections, prosthetic mobilization) that involve the labeling, using radioactive molecules, of autologous cells, whose localization can then be detected using standard diagnostic tools (nuclear medicine instrumentation). However, the use of radio-emitting molecules limits the application of these procedures: the signal remains detectable only for a limited period of time, which depends on the half-life of the radionuclide used. Therefore, if applied in the context of cell-mediated treatments, these imaging procedures would be able to provide only initial information about the accuracy of the transplant and the early general distribution of the cells. For this reason, a more stable label needs to be identified and tested, also for clinical applications, with the aim of allowing long-term imaging of cell localization. Among the already approved contrast agents and imaging procedures, small paramagnetic nanoparticles able to produce an MRI signal (iron- and gadolinium-based agents) have already been tested for cell imaging applications 43 , 44 , 45 , 46 , 47 and, in our opinion, are destined, in the near future, to prove extremely important in the evaluation of several cell-mediated treatments.
Herein, we demonstrated the possibility of efficiently and safely labeling murine adult NSCs with paramagnetic nanoparticles. Specifically, we used SPIOs (Endorem®) as contrast agents and PrS as a carrier, since both are approved for clinical use. Dose-response and incubation time studies allowed us to optimize the procedure in order to obtain homogeneous and efficient cell labeling. The best labeling conditions for adult NSCs was the incubation with 200 μg of iron/ml of culture medium for 24 hours in presence of PrS (ratio Fe/PrS 1:0.025). This protocol represented the best possible compromise between degree of labeling and cell viability, while also producing an iron load per cell sufficient for providing a signal detectable by in vivo MRI. Such iron-labeled NSCs showed no changes in phenotypic differentiation as shown by qualitative and quantitative estimates. Thus SPIOs labeling did not negatively influence their viability and cellular properties. Through their differentiation in astrocytes, oligodendrocytes and neurons. On the basis of these results, we can conclude that clinically approved reagents can be used to label NSCs with SPIOs without influencing cell-specific features, and allow their in vivo imaging after administration.
The main problem in setting up of an imaging protocol for the evaluation of cell-mediated treatments is the need to visualize even small amounts of cells within the target organ. Gradient echo (FLASH) MRI sequences were initially chosen because they show a higher sensitivity to susceptibility effects, such as that attributable to the presence of iron, while providing sufficient anatomical detail to allow identification of the lesion site.
Several reports have described MRI of different stem cell types injected intra-spinally both in the rat and in the murine model of SCI. In particular, Sykovà et al. reported that mesenchymal stem cells (MSCs), after intravenous injection in the rat SCI model, could be visualized only by means of ex vivo MRI because of the presence, in the living model, of breathing-derived noise in the thoracic region where the spinal lesion is located 48 , 49. Another recent paper by Gonzalez-Lara et al. showed for the first time the possibility of performing MRI in vivo in spinal cord injured mice. However, the technique could reveal only intra-spinally injected iron-loaded MSCs. This paper also describes a very interesting observation that is how the hypointense region changed in volume over time depending on the condition of the applied cells that were alive or dead 50.
To our knowledge, our study is the first to use in vivo MRI to evaluate the localization of NSCs at the injured spinal cord following their intravenous injection in the mouse tail vein. In spite of difficulties in finding the right plane to image the entire spinal cord due to scoliosis caused by laminectomy and the traumatic injury to the cord, MRI evaluation was able to show the accumulation of NSCs within the target area with high sensitivity and provided images containing anatomical information important for the evaluation of cell localization. In particular, RARE images, being strongly T2-weighted, highlighted the shrinkage of the spinal cord within 3 weeks after the injury, and the consequent signal increase due to the changes in the liquor/spinal cord ratio at the lesion site. This element could be used in the future to assess the efficacy of cell-mediated treatment in reducing the consequences of damage. MRI showed high homogeneity of the hypointense signal in all the intravenously injected animals, demonstrating that the procedure was highly consistent, despite the complexity of the experimental model.
As regards to the assessment of the hypointense volume over time, future in vivo examinations will seek to quantify the extent of the iron signal in order to evaluate whether there is a correlation between functional recovery and accumulation of labeled cells at the lesion site over time. In this paper, our aim was to describe the real possibility of performing NSC imaging in a cell-mediated treatment protocol using clinically approved reagents and a procedure readily applicable to humans. Future studies will focus on relating functional recovery after treatment to MRI data even though it has been demonstrated in this paper how SPIO labelling procedures do not negatively influence the NSC promoted enhancement of locomotor recovery. However, cell-mediated treatment efficacy needs to be studied further to assess the relationship between the amount of therapeutic cell present at the injury site and motor recovery.
The choice of MRI as diagnostic imaging tool is extremely significant. After all, this technique is routinely applied, involves no radiation exposure risks and lends itself to longitudinal studies. It is also capable of providing images containing different data such as the hypointense signal due to iron-labeled cell localization, the hyperintense signal due to the change in the balance between spinal cord and liquor at the lesion site, and anatomical information describing the lesion structure. In addition, given its ability to detect the presence of SCs and to describe anatomical features of lesions, it emerges as the ideal technique for evaluating whether the presence of SCs can reduce the consequences of injury.
The data reported in this paper demonstrate the value of our model for the in vivo study of key variables in the efficacy evaluation of a cell-mediated therapy, such as the ability of the transplanted cells to migrate, and localize at the target, which is the first step in an efficient therapeutic protocol, and their persistence over time.
ACKNOWLEDGEMENTS
We wish to thank Dr E. Pesenti (Nerviano Medical Sciences) for the use of MRI system and Dr. Laura Madaschi (Depatment of Health Sciences, University of Milan) for technical support. D.C. is supported by a fellowship from the Doctorate School of Molecular Medicine, University of Milan; T.G. is supported by a fellowship from the Doctorate School of Biochemical, Nutritional and Metabolic Sciences.
The authors are grateful to Ms Catherine Wrenn for her advice and skilful editorial support. This study was supported by a Cariplo Foundation grant (2007.5281), Neurogel en Marche Foundation (to Alfredo Gorio).
The authors declare that they have no conflict of interest.
References
- Gorio A, Gokmen N, Erbayraktar S, Yilmaz O, Madaschi L, Cichetti C, Di Giulio AM, Vardar E, Cerami A, and Brines M. Recombinant human erythropoietin counteracts secondary injury and markedly enhances neurological recovery from experimental spinal cord trauma. Proc Natl Acad Sci U S A 2002; 99: 9450-9455. (back)
- Gorio A, Madaschi L, Di Stefano B, Carelli S, Di Giulio AM, De Biasi S, Coleman T, Cerami A, and Brines M. Methylprednisolone neutralizes the beneficial effects of erythropoietin in experimental spinal cord injury. Proc Natl Acad Sci U S A 2005; 102: 16379-16384. (back)
- Kwon BK, Tetzlaff W, Grauer JN, Beiner J, Vaccaro AR. Pathophysiology and pharmacologic treatment of acute spinal cord injury. Spine J 2004;4(4):451-464.) , ((Barbon A, Fumagalli F, Caracciolo L, Madaschi L, Lesma E, Mora C, Carelli S, Slotkin TA, Racagni G, Di Giulio AM, Gorio A, Barlati S. Acute spinal cord injury persistently reduces R/G RNA editing of AMPA receptors. J Neurochem. 2010; 114(2): 397-407. (back)
- Pertici V, Amendola J, Laurin J, Gigmes D, Madaschi L, Carelli S, Marqueste T, Gorio A, Decherchi P. The use of poly(N-[2-hydroxypropyl]-methacrylamide) hydrogel to repair a T10 spinal cord hemisection in rat: a behavioural, electrophysiological and anatomical examination. ASN Neuro 2013; 5(2): 149-166. (back)
- Caracciolo L, Fumagalli F, Carelli S, Madaschi L, La Via L, Bonini D, Fiorentini C, Barlati S, Gorio A, Barbon A. Kainate receptor RNA editing is markedly altered by acute spinal cord injury. J Mol Neurosci 2013; 51(3): 903-910. (back)
- Bottai D, Madaschi L, Di Giulio AM, Gorio A Viability-dependent promoting action of adult neural precursor in spinal cord injury. Mol Med 2008; 14(9-10): 634-644. (back)
- Bottai D, Cigognini D, Nicora E, Moro M, Grimoldi MG, Adami R, Abrignani S, Marconi AM, Di Giulio AM, Gorio A. Third trimester amniotic fluid cells with the capacity to develop neural phenotypes and with heterogeneity among sub-populations. Restor Neurol Neurosci 2011; 30(1): 55-68. (back)
- Horn KP, Busch SA, Hawthorne AL, van Rooijen N, Silver J. Another barrier to regeneration in the CNS: activated macrophages induce extensive retraction of dystrophic axons through direct physical interactions. J Neurosci 2008; 28: 9330-9341. (back)
- Bottai D, Scesa G, Cigognini D, Adami R, Nicora E, Abrignani S, Di Giulio AM, Gorio A. Third trimester NG2-positive amniotic fluid cells are effective in improving repair in spinal cord injury. Exp Neurol pii: 2014; S0014-4886(14)00028-4. (back)
- Bottai D, Madaschi L, Di Giulio AM, Gorio A Viability-dependent promoting action of adult neural precursor in spinal cord injury. Mol Med 2008; 14(9-10): 634-664. (back)
- Lecchi M, Ottobrini L, Martelli C, Del Sole A, Lucignani G. Instrumentation and probes for molecular and cellular imaging. Q J Nucl Med Mol Imaging 2007; 51(2): 111-126. (back)
- Yang Y, Schumacher A, Yang Y, Liu J, Shi X, Hill WD, Hu TCC. Monitoring bone marrow-originated mesenchymal stem cell traffic to myocardial infarction sites using magnetic resonance imaging. Magn Reson Med 2011; 65(5): 1430-1436. (back)
- Chien LY, Hsiao JK, Hsu SC, Yao M, Lu CW, Liu HM, Chen YC, Yang CS, Huang DM. In vivo magnetic resonance imaging of cell tropsim, trafficking mechanism, and therapeutic impact of human mesenchymal stem cells in a murine glioma model. Biomaterial 2011; 32(12): 3275-3284. (back)
- Xu Q, Zhang HT, Liu K, Rao JH, Liu XM, Wu L, Xu BN. In vitro and in vivo magnetic resonance tracking of sinerem-labeled human umbilical mesenchymalstromal cell-derived Schwann cells. Cell Mol Neurobiol 2011; 31(3): 365-375. (back)
- Meng X, Seton HC, Lu LT, Prior IA, Thanh NT, Song B. Magnetic CoPt nanoparticles as MRI contrast agent for transplanted neural stem cells detection. Nanoscale 2011; 3: 977-984. (back)
- Jiang H, Cheng Z, Tian M, Zhang H. In vivo imaging of embryonic stem cell therapy. Eur J Nucl Med Mol Imaging 2011; 38(4): 774-784. (back)
- Neri M, Maderna C, Cavazzin C, et al. Efficient in vitro labeling of human neural precursor cells with superparamagnetic iron oxide particles: relevance for in vivo cell tracking. Stem Cells 2008; 26: 505-516. (back)
- Daldrup-Link HE, Rudelius M, Oostendorp RAJ, et al. Targeting of hematopoietic progenitor cells with MR contrast agents. Radiology 2003; 228: 760-767. (back)
- Frank JA, Zywicke H, Jordan EK, et al. Magentic intracellular labeling of mammalian cells by combining (FDA-approved) superparamegnetic iron oxide MR contrast agents and commonly used transfection agents. Acad Radiol 2002; 9: S484-448. (back)
- Bulte JWM, Kraitchman DL. Iron oxide MR contrast agents for molecular and cellular imaging. NMR Biomed 2004; 17: 484-499. (back)
- Dunning MD, Lakatos A, Loizou L, et al. Superparamagnetic iron oxide labeled Schwann cells and olfactory ensheathing cells can be traced in vivo by magnetic resonance imaging and retain functional properties after transplantation into the CNS. J Neurosci 2004; 24: 9799-9810. (back)
- Zhang Y, Ruel M, Beanlands RS, deKemp RA, Suuronen EJ, DaSilva JN. Tracking stem cell therapy in the myocardium: applications of positron emission tomography. Curr Pharm Des 2008; 14(36): 3835-3853. (back)
- Doyle B, Kemp BJ, Chareonthaitawee P, Reed C, Schmeckpeper J, Sorajja P, Russell S, Araoz P, Riederer SJ, Caplice NM. Dynamic tracking during intracoronary injection of 18F-FDG-labeled progenitor cell therapy for acute myocardial infarction. J Nucl Med 2007; 48(10): 1708-1714. (back)
- Jendelová P, Herynek V, Urdzíková L, Glogarová K, Kroupová J, Andersson B, Bryja V, Burian M, Hájek M, Syková E. Magnetic resonance tracking of transplanted bone marrow and embryonic stem cells labeled by iron oxide nanoparticles in rat brain and spinal cord. J Neurosci Res 2004; 76(2): 232-243. (back)
- Lee IH, Bulte JWM, Schweinhardt P, Douglas T, Trifunovski A, Hofstetter C, Olson L, and Spengera C. In vivo magnetic resonance tracking of olfactory ensheathing glia grafted into the rat spinal cord. Exp Neurol 2004; 187: 509-516. (back)
- Gonzalez-Lara LE, Xu X, Hofstetrova K, Pniak A, Chen Y, McFadden CD, Martinez-Santiesteban FM, Rutt BK, Brown A, Foster PJ. The use of cellular magnetic resonance imaging to track the fate of iron-labeled multipotent stromal cells after direct transplantation in a mouse model of spinal cord injury. Mol Imaging Biol 2010; 13(4): 702-711. (back)
- Iwanami A, Kaneko S, Nakamura M, Kanemura Y, Mori H, Kobayashi S, M. Yamasaki, Momoshima S, Ishii H, Ando K, Tanioka Y, Tamaoki N, Nomura T, Toyama Y, and Okano H. Transplantation of human neural stem cells for spinal cord injury in primates. J Neurosci Res 2005; 80:182-190. (back)
- Gritti A, Bonfanti L, Doetsch F, Caille I, Alvarez-Buylla A, Lim DA, Galli R, Verdugo JM, Herrera DG, Vescovi AL. Multipotent neural stem cells reside into the rostral extension and olfactory bulb of adult rodents. J Neurosci 2002; 22(2): 437-445. (back)
- Gritti A, Parati EA, Cova L, Frolichsthal P, Galli R, Wanke E, Faravelli L, Morassutti DJ, Roisen F, Nickel DD, Vescovi AL. Multipotential stem cells from the adult mouse brain proliferate and self-renew in response to basic fibroblast growth factor. J Neurosci 1996; 16(3): 1091-1100. (back)
- Marfia G, Madaschi L, Marra F, Menarini M, Bottai D, Formenti A, Bellardita C, Di Giulio AM, Carelli S, Gorio A. Adult neural precursors isolated from post mortem brain yield mostly neurons: an erythropoietin-dependent process. Neurobiol Dis 2011; 43(1): 86-98. (back)
- Boutry S, Brunin S, Mahieu I, Laurent S, Vander Elst L, Muller RN. Magnetic labeling of non-phagocy tic adherent cells with iron oxide nanoparticles: a comprehensive study. Contrast Media Mol Imaging 2008; 3: 223-232. (back)
- Montet-Abou K, Montet X, Weissleder R, Josephson L Cell internalization of magnetic nanoparticles using transfection agents. Mol Imaging 2007; 6(1): 1-9 (back)
- Marfia G, Madaschi L, Marra F, Menarini M, Bottai D, Formenti A, Bellardita C, Di Giulio AM, Carelli S, Gorio A. Adult neural precursors isolated from post mortem brain yield mostly neurons: an erythropoietin-dependent process. Neurobiol Dis 2011; 43(1): 86-98. (back)
- Gorio A, Gokmen N, Erbayraktar S, Yilmaz O, Madaschi L, Cichetti C, Di Giulio AM, Vardar E, Cerami A, and Brines M. Recombinant human erythropoietin counteracts secondary injury and markedly enhances neurological recovery from experimental spinal cord trauma. Proc Natl Acad Sci U S A 2002; 99: 9450-9455. (back)
- Gorio A, Madaschi L, Di Stefano B, Carelli S, Di Giulio AM, De Biasi S, Coleman T, Cerami A, and Brines M. Methylprednisolone neutralizes the beneficial effects of erythropoietin in experimental spinal cord injury. Proc Natl Acad Sci U S A 2005; 102: 16379-16384. (back)
- Gorio A, Gokmen N, Erbayraktar S, Yilmaz O, Madaschi L, Cichetti C, Di Giulio AM, Vardar E, Cerami A, Brines M. Recombinant human erythropoietin counteracts secondary injury and markedly enhances neurological recovery from experimental spinal cord trauma. Proc Natl Acad Sci U S A 2002; 99: 9450-9455. (back)
- Gorio A, Madaschi L, Di Stefano B, Carelli S, Di Giulio AM, De Biasi S, Coleman T, Cerami A, and Brines M. Methylprednisolone neutralizes the beneficial effects of erythropoietin in experimental spinal cord injury. Proc Natl Acad Sci U S A 2005; 102: 16379-16384. (back)
- Boutry S, Brunin S, Mahieu I, Laurent S, Vander Elst L, Muller RN. Magnetic labeling of non-phagocy tic adherent cells with iron oxide nanoparticles: a comprehensive study. Contrast Media Mol Imaging 2008; 3: 223-232. (back)
- Bottai D, Madaschi L, Di Giulio AM, Gorio A. Viability-dependent promoting action of adult neural precursor in spinal cord injury. Mol Med 2008; 14(9-10): 634-664. (back)
- Bottai D, Madaschi L, Di Giulio AM, Gorio A. Viability-dependent promoting action of adult neural precursor in spinal cord injury. Mol Med 2008; 14(9-10): 634-664. (back)
- Lecchi M, Ottobrini L, Martelli C, Del Sole A, Lucignani G. Instrumentation and probes for molecular and cellular imaging. Q J Nucl Med Mol Imaging 2007; 51(2): 111-126. (back)
- Lecchi M, Ottobrini L, Martelli C, Del Sole A, Lucignani G. Instrumentation and probes for molecular and cellular imaging. Q J Nucl Med Mol Imaging 2007; 51(2): 111-126. (back)
- Bulte JWM, Kraitchman DL Iron oxide MR contrast agents for molecular and cellular imaging. NMR Biomed 2004; 17: 484-499. (back)
- Libani IV, Lucignani G, Gianelli U, Degrassi A, Russo M, Bosari S, Clerici M, Ottobrini L. Labeling protocols for in vivo tracking of human skeletal muscle cells (HSkMCs) by magnetic resonance and bioluminescence imaging. Mol Imaging Biol 2011; 14(1): 47-59. (back)
- Kraitchman DL, Kedziorek DA, Bulte JW. MR imaging of transplanted stem cells in myocardial infarction. Methods Mol Biol 2011; 680: 141-152. (back)
- Son KR, Chung SY, Kim HC, Kim HS, Choi SH, Lee JM, Moon WK. MRI of magnetically labeled mesenchymal stem cells in hepatic failure model. World J Gastroenterol 2010; 16(44): 5611-5615. (back)
- Sykovà E, Jendelovà P. Magnetic Resonance Tracking of implanted Adult and Embryonic stem cells in injured brain and spinal cord. Ann NY Acad Sci 2005; 1049: 146-160. (back)
- Bottai D, Madaschi L, Di Giulio AM, Gorio A. Viability-dependent promoting action of adult neural precursor in spinal cord injury. Mol Med 2008; 14(9-10): 634-664. (back)
- Sykovà E, Jendelovà P. Magnetic Resonance Tracking of implanted Adult and Embryonic stem cells in injured brain and spinal cord. Ann NY Acad Sci 2005; 1049: 146-160. (back)
- Libani IV, Lucignani G, Gianelli U, Degrassi A, Russo M, Bosari S, Clerici M, Ottobrini L. Labeling protocols for in vivo tracking of human skeletal muscle cells (HSkMCs) by magnetic resonance and bioluminescence imaging. Mol Imaging Biol 2011; 14(1): 47-59. (back)
To cite this article
Magnetic Resonance Imaging of Stem Cell Transplantation in Injured Mouse Spinal Cord
CellR4 2014; 2 (3): e963
Publication History
Published online: 31 May 2014

This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.